第一节 建立转基因动物的方法
1.目的基因的设计
在进行转基因动物研究时,首先要针对实验目的设计待转移的目的基因,如随机整合型基因或基因敲除(gene knockout)型载体基因。随机整合型基因可来自于同一基因,也可来自于不同基因(拼接基因)。从结构上看,随机整合型基因可分为结构基因与调控序列两部分。若来自于同一个基因,那么该基因必须完整,包括5'上游启动子区和3'下游的加尾信号(polyA signal)区等,可从基因组文库中分离完整的基因。拼接基因应用更为广泛,可通过选择不同的启动子来控制外源基因表达的组织特异性。进行基因剔除转基因动物研究时,首先要设计与待剔除基因的同源性的载体基因,包括正负选择标志基因,通过同源重组(同源取代或同源插入)而使目的基因失活,从而建立缺基因动物。在进行基因设计时,还应考虑转基因动物的遗传背景和设计相应的检测方法,必要时设计一个报道基因(reporter gene)。
2.载体的选择
运用转基因动物研究基因表达,特别是在研究遗传性疾病和进行基因治疗时,选择合适的基因载体是十分必要的。载体主要用于未克隆基因的扩增和选择,有关实验表明,载体有DNA序列可以干扰外源基因的表达,其原因可能与DNA的甲基化有关。最早用于哺乳类细胞表达载体的病毒是转化型DNA病毒,如SV[XB]40[/XB]和腺病毒。这些载体的主要缺点是接受外源DNA容量太小,只能携带约7~8kb的外源序列,因此这类载体不可能表达其他许多与疾病有关的功能基因。与其他载体相比,从鸟类和鼠类中获得的逆转录病毒是目前应用广泛的一类最有前途的载体。这类病毒为失去致病能力但仍有感染能力的缺陷病毒,如通常使用的禽白血病病毒和罗氏肉瘤病毒。这类病毒载体能有效地把外源基因导入哺乳动物靶细胞,其整合方式在结构和功能上较稳定,功能基因插入和表达很容易。为避免逆转录病毒启动子干扰和携入诱变的可能性,许多研究者还构建出一类已被剪除了自身启动子和增强子序列的逆转录病毒。这类失活载体只表达由基因携带的启动子起始的载体编码序列,但这种载体因其滴度低而使应用受到限制。近来有报道指出,利用带有部分缺失失活的新霉素抗性基因(neo[XB]r[/XB])的缺陷型整合逆转录病毒载体感染大鼠细胞,使整合到细胞中的neo[XB]r[/XB]缺陷基因功能得到恢复。非整合型病毒载体,如HSV载体已引起人们极大关注,这类病毒基因组较大(150kb),它们具有比其他已知载体接受更大外源序列的能力,故能够转移和表达较大的基因。
3.背景及种系的选择
由于不同种系的动物对不同疾病的易感性不同,在进行转基因动物研究时要考虑遗传背景及种系的问题。许多实验室在随机杂交背景下建立了转基因动物,同近亲交配的转基因动物相比,这一背景下产生的胚胎不但数量多,而且健康。尽管如此,随机交配下的碱基分离可能影响转基因动物的表型,降低了一个已知转基因动物的作用。在混合遗传背景下,碱基分离也直接影响基因动物的表达。已发现一种能够直接抑制了转基因的表达的种系特异性修饰因子(SSM-1)。在C57BL/6J鼠系中,SSM-1碱基分离直接抑制了转基因的表达。相反,在DBA/2JSSM-1鼠中可交配的DBA/2J小鼠,可以消除SSM-1碱基分离产生变异的转基因表达。
迄今为止,已相继报道了小鼠、大鼠、兔、猪、牛和羊等转基因的动物。目前,研究者均倾向于使用小鼠作转基因的动物。最初的转基因动物就是在小鼠体内完成的,已确定了小鼠的连锁图。尽管由于小鼠与人脂蛋白系统有很大差异,如小鼠缺乏胆固醇酯转移蛋白(CETP)、载脂蛋白(a)[Apo(a)]和脂蛋白(a)[Lp(a)],其多数胆固醇通过HDL而不像人那样通过LDL进行运输,但由于小鼠易于饲养管理,生长周期短,容易获得已经定性的近交交配鼠和遗传突变体,并且易于进行快速连锁分析,其作为研究动物也较为经济实惠,故常被用作建立转基因动物模型。但小鼠对AS具有抗性,为使小鼠产生AS,必需喂养非生理性食物,包括1.25%的胆固醇(为人类饮食的10~20倍),15%的脂肪和0.5%的胆酸(非正常食物组分)。严格地讲,这种饲料是具有毒性的,但可使小鼠的非高密度脂蛋白胆固醇(HDL-C)增至200~300mg/ml,4~5个月即可形成AS。
4.转基因方法
进行转基因动物研究的基因转移方法有多种,如早期的畸胎癌细胞(teratocarcinoma cell,TCC)植入法、逆转录病毒感染法、显微镜注射法以及近几年新出现的方法如电转移法,精子载体法,胚胎干细胞(embryonal sterm cell,ES细胞)法、基因直接导入法等。其中较常用的三种方法有显微注射法、逆转录病毒感染法和胚胎干细胞法(见图23-1)。
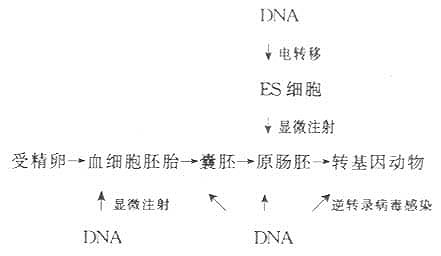
图23-1 三种常用的转基因方法
显微注射法是直接将重组DNA分子以微注射的方式导入单细胞卵的原核中,再将它植入假孕母鼠。大约20%的微注射胚胎能够将外源基因整合到染色体基因上组上;大多数的转基因的动物能够将整合的基因传给后代,建立起转基因鼠系。逆转录病毒感染法是用高滴度的、携带外源基因的重组逆转录病毒感染发育早期的胚胎,再将感染病毒后的胚胎植入假孕母鼠,以产生转基因的动物。逆转录病毒的整合并不影响宿主DNA序列的重排,感染的复数容易调整,每个卵大约有10次整合的机会。病毒染色体还能提供一个TAG分子,加速覆盖部位的迅速克隆。这些特征使逆转录病毒转基因法用于研究随机突变。从ES细胞到转基因动物是从哺乳动物胚胎中分离出ES细胞,通过转导或传染的方法,将外源基因转入ES细胞,再以微注射的方式将其植入鼠的胚胎。这种方法能够将外源基因定位导入靶细胞染色体上某一特定部位,或使某一基因发生定点突变。
上述三种方法各具有优缺点。显微注射法相对整合率较高(1%~10%),操作简便,易于获得成功。但外源基因通常以多拷贝串联的形式随机整合于受体基因组中,这种异常排列可能妨碍其表达的正常调节。逆转录病毒感染法的外源基因通常是单位点、单一拷贝整合,整合通常发生在逆转录病毒的长末端重复区(LTR),从而保证了外源基因结构的完整性。但整合效率较低,而且操作比较繁琐,能被导入的基因大小有一定限制(通常为10kb左右)。ES细胞法多只能建立嵌合体动物,如果注射有外源基因的ES细胞在胚胎内的发育过程中嵌合到生殖腺,则所建立的嵌合体动物不能把外源性基因传给后代。但通过ES细胞法可以进行基因打靶(gene targeting)或基因剔除产生缺基因动物,这是目前建立某些疾病动物模型的一种新的有效途径。
5.转基因动物模型的建立
新出生的转基因动物通过点杂交、聚合酶链反应(PCR)和免疫印迹杂交(Southern Northern blotting)等方法,先筛选出有外源基因整合的阳性动物,传代后分别在mRNA和蛋白水平上检测外源基因在转基因动物中的表达情况,对外源基因表达产物的生物活性和生化性质进行鉴定,以及检查转基因动物的生理功能和是否出现某些疾病病症状的表型等。进一步培育和筛选出转基因动物的纯合子,或/和某些相关种系的动物杂交,从而建立某种疾病的转基因动物模型。
利用转基因技术培育人类疾病动物模型比用其他方法更加优越,它可在动物原来遗传背景的基因上,通过改变某种基因的表达水平而实现。这种模型模拟动物症状单一,接近于病人症状,产生这些疾病症状的原因是外源基因的转入。转基因动物模型可按照人们的愿望进行设计和培育,其建立过程的本身就可进行疾病机理的研究。由于转基因动物的发育过程中又引入了时间和空间的因素,这样就建立了一个立体的实验动物体系,不但能从动物整体水平的组织器官水平上进行研究,而且还可以深入到细胞水平和分子水平,为发病机理、药物筛选和临床医学研究提供了比较理想的实验动物体系。自从1980年第一个转基因动物问世以来,经过科学家们的不懈努力,已相继建立了高脂血症(HLP)、AS、高血压、低血压等心血管疾病的转基因动物模型。其中应用最广的是各种转基因小鼠(TGM)模型。