第七节 计划生育研究中实验动物的选择和应用
一、常用实验动物的生殖功能、结构特点
实行计划生育是我国的一项基本国策。到本世纪末,力争把我国人中控制在12亿以内,此项任务十分艰巨。如果人口控制不住,就会影响经济振兴,影响现代化,甚至影响社会的安定。广大人民群众迫切需要更多、更安全的、使用方便的节育措施。为了寻找更多的有效措施来控制人类的生育,保证避孕药物或其他节育措施的有效和安全,必须首先在实验动物身上进行实验研究。因此,保证实验动物的质量和了解实验动物生殖和发育等特点,是十分重要。
(一)常用实验动物的繁殖周期特点
计划生育研究中常用的实验动物有小鼠、大鼠、豚鼠、兔狗、恒河猴等,各种动物的性周期、排卵时间及妊娠期都各不相同。如狗是季节性单发情,猫季节性多次发情,而鼠、豚鼠全年都有动情周期的循环,灵长类有月经周期,但只有一定季节才排卵(见下页表10-37)
(二)常用实验动物排卵和性周期特点
哺乳动物的排卵可分为两种类型,一般动物是按一定的周期进行排卵的,这类动物不交配也可正常排卵,此称为自发排卵或非刺激性排卵;而兔子、猫、雪貂(白鼬)、水貂等,只有经过交配的刺激才能进行排卵,此称为诱发排卵,也称刺激性排卵或反射性排卵。例如兔子的卵巢几乎连续不断地产生卵子,但成熟的卵泡只有经过雄兔交配后才能排卵。这是因为雌兔受到雄兔爬跨时,垂体受到兴奋而引起的刺激,这种刺激出现在交配后1小时,此时垂体释放排卵素(黄体生物素),在促排卵素的影响下大约在交配后10~12小时发生排卵。如果没有交配刺激,成熟的卵泡自行退化,经10~16小时逐渐被吸收,而新的卵泡又开始成熟。
雌性哺乳动物的性周期可分为三型
第一型包括狗、马、牛以及其他哺乳动物的性周期。该型性周期又可分为前半期和后半期。前半期由雌激素所控制,雌激素的效果逐渐变强,至发情期达到最高程度。这个时期,卵巢内卵泡成熟,随后便引起排卵,随着排卵,雌激素的作用逐渐减弱,以后即转入以孕激素所控制的后半期。如果没有妊娠,黄体退化,则又转为由雌激素所控制的新的性周期。总之,在这一性周期中,雌激素作用的期间和孕激素作用的期间是相继出现的,在两期之间排卵。第一型是基本型。
第二型包括小白鼠、大白鼠、田鼠类、土拔鼠和沙鼠的性周期。这类动物在发情期排卵,也形成黄体,但生成的黄体不能分泌孕激素并立即退化,即没有第一型的后半期,所以性周期是短的。如果妊娠了,黄体继续存在,称为妊娠黄体,能分泌黄体酮。
第三型包括兔子、猫等的性周期,这类动物如果不交配就不引起排卵,也不形成黄体,成熟的卵泡逐渐退化消失,随后又有新的卵泡成熟,并不断反复进行更新,这种情况下的性周期约为15~16天(成熟卵泡的寿命为13天,卵泡退化约1~2天);如果进行交配,则引起排卵,产生黄体,分泌孕激素;若没有受精,则所形成的假妊娠黄体约经20天退化。
表10-37 实验动物繁殖周期
| 动物种类 | 第一次交配年龄 | 发情周期(天) | 发情持续期 | 排卵时间 | 受精卵进入子宫时间 | 植入开始时间 | 妊娠期 |
| 狗 | 1(1/2)~2岁雄狗2岁 | 发情期间隔4~8个月,春秋以品种不同而不同 | 9(4~13)天 | 发情第3或4天 | 交配后5~6天 | 交配后15天 | 60天(58~63) |
| 猫 | 6~8月龄 | 季节性强,春秋季节多次发情15~28 | 4(3~10)天 | 诱发排卵,交配后27小时 | 排卵后4天 | 交配后13天 | 63天(52~69) |
| 兔 | 4~5月龄 | 多次发情 | 时间界限不明显 | 诱发排卵,交配后10.5小时 | 交配后4天 | 交配后7~8天 | 32天(29~36) |
| 大白鼠 | 8~10月龄 | 4.5(4~5) | 14小时 | 发情开始后10小时 | 交配后3~4天 | 交配后5~6天 | 22天(19~23) |
| 小白鼠 | 6~8月龄 | 5(4~7) | 6~24小时 | 发情开始后2~3小时 | 交配后3~4天 | 交配后4~5天 | 19-21天 |
| 豚鼠 | 雄性6月龄雌性5月龄 | 16.5(12~18) | 8小时 | 发情开始后10小时 | 交配后3~4天 | 交配后5~7天 | 70天(69~75) |
| 恒河猴 | 第3~5次性周期后才可进行交配 | 月经周期28(23~33) | 时间界限不明显一般为4~6小时 | 月经周期9~20天接近发情结束时 | 交配后4天 | 交配后9天 | 164天(156~180) |
实验动物性季节(繁殖季节)差异较大,兔、小白鼠、大白鼠、猪等一年四季都有性的活动(妊娠时除外),这类动物称为“常年多次发情(或常年发情动物)”,其繁殖力很强。狗的发情有一定的季节性,雌狗一年内发情两次,多在春秋两季,每次发情持续14~21天,每次发情的间隔期约7个月。猫的繁殖季节,北半球是3~8月分,南半球是9月到次年的2月份。欧洲田鼠的繁殖季节是4~8月份。羊的繁殖季节在秋季(7~10月份),羊也是“季节性多次发情”。
实验动物性周期一般分为四期:发情前期,发情期、发情后期和发情间期(休情期)。有的还将发情后期分为二期(后情一期和二期)。
小鼠动情周期一般分为五期(其中动情后分为二期),约120~144小时。大鼠动情周期为96~120小时。
哺乳类雌性动物在性成熟后出现动情周期变化,其中啮齿类动物在动情周期不同阶段,阴道粘膜发生比较典型的变化。因次,根据阴道涂片法的细胞学改变,可以推断卵巢功能的周期变化。方法是用圆头滴管吸少许生理盐不滴入阴道口,轻轻冲吸一点阴道液体作涂片,待涂片干燥后,用10%福尔马林或95%酒精固定,经苏木精一伊红染色后作镜检(表10-38)。
表10-38 动情周期阴道涂片的细胞变化特点
| 阶 段 | 经过时间(小时) | 卵巢变化 | 细 胞 变 化 特 点 | |
| 小鼠 | 大鼠 | |||
| 动情前期(P)(Proestrus Stage) | 10 | 17~21 | 卵泡加速生长 | 全部是有核上皮细胞,偶有少量角化细胞 |
| 动情期(E)(Estrus Stage) | 42 | 9~15 | 卵胞成熟排卵 | 全部是无核角化细胞或间有少量上皮细胞 |
| 动情后期(M)(Metestrus Stage) | 12 | 10~14 | 黄体生成 | 白细胞、角化细胞、有核上皮细胞均有 |
| 动情间期(DI)(Diestrus Stage) | 48~72 | 60~70 | 黄体退化 | 大量白细胞及少量上皮细胞和粘膜 |
(三)常用实验动物精子和卵子的运输特点
1.精子的运输哺乳动物精子从阴道到输卵管是很快的。马、猪和大鼠在交配后很短时间,子宫腔内就看到了精子。这些动物由于在交配时宫颈口舒张,所以精子直接射到子宫腔内。兔精子进入子大约需要1~3小时。
精子获能是一个生理过程,即精子获得穿透新排出卵的透明带的能力。这个过程对一些动物的受精作用是必需的。兔精子获能需要6小时,大鼠2~3小时,羊约1.5小时,雪貂大约3.5小时。实验证明兔、大鼠、小鼠、地鼠、羊和雪貂精子的获能是必需的。
哺乳动物精子在进入雌性生殖道以后,对卵的受精能力局限于很短一段时间。各种动物在雌性生殖道内有生育能力的时间有很大变化(表10-39)。
表10-39 精子在雌性生殖道内的生育力和活动力的时间
| 实验动物 | 生育力最长时间(小时) | 活动力最长时间(小时) |
| 小 鼠 | 6 | 13 |
| 大 鼠 | 14 | 17 |
| 豚 鼠 | 21~22 | 41 |
| 兔 | 30~32 | - |
| 雪 貂 | 36~48;126 | - |
| 牛 | 28~50 | 96 |
| 羊 | 30~48 | 48 |
| 马 | 144 | 144 |
| 蝙 蝠 | 135天 | 159天 |
在整个动情期,小白鼠子宫为液体充满而扩张,子宫壁产生蠕动,可以推动精子随液体进入输卵管,在输卵管内,精子移动较慢,约在交配后15分钟到1小时之内,始能到达输卵管壶腹部。精子在输卵管内保持受精力不超过6小时,但活动力可长达13.5小时。精子要经过在子宫或输卵管中液体的处理后才能获得受精能力。
2.卵的运输当卵泡破裂时,有一部分卵泡液随之流出,卵被排到围卵巢腔内,所以不发生漏到腹腔内的现象。当动情期时,围卵腔变大并产生一种液体,能把卵带到输卵管伞部,加之漏斗部的绒毛活动和输卵管的收缩帮助卵运输到输卵管下段。
卵子的大小并不取决于动物的个体,而几乎完全取决于所积累的卵黄含量的多少。各种常用实验动物的卵子直径(μm)如下:
鼷鼠 75~87.8 狗 135~145 猫 120~130
小白鼠 70~75 豚鼠 75~85 水貂 107
大白鼠 70~75 兔 120~120 猴 125~143
卵由卵巢滤泡运输到子宫腔的时间约3~4天,常用实验动物输卵管内卵的运输情况和受精时间见表10-40和10-41。
表10-40 卵子经过输卵管的时间
| 动 物 | 出发点 | 卵在输卵管的时间(天) | 进入子宫时间 |
| 人 | 排 卵 | 3 | 12个细胞 |
| 猴 | 排 卵 | 4 | 16个细胞 |
| 兔 | 交 配 | 3-4 | 桑椹早期总胚泡后期 |
| 仓 鼠 | 交 配 | 2.5 | 4-8个细胞 |
| 豚 鼠 | 交 配 | 4 | 4-8个细胞 |
| 大 鼠 | 交 配 | 3.5-5 | 桑椹早期至胚泡后期 |
| 小 鼠 | 交 配 | 3 | 桑椹早期至胚泡后期 |
| 牛 | 排 卵 | 3-4 | 8-16个细胞 |
| 猪 | 交 配 | 4-5 | 3-4个细胞 |
| 羊 | 交 配 | 3-4 | - |
10-41 哺乳动物卵的受精时间
| 动 物 | 受精时间(小时) |
| 人 | 6-24 |
| 猴 | 23 |
| 兔 | 6-8 |
| 仓 鼠 | 5 |
| 豚 鼠 | 20 |
| 大 鼠 | 12 |
| 小 鼠 | 8 |
| 雪 貂 | 30 |
| 牛 | 18-20 |
| 绵 羊 | 24 |
(四)常用实验动物着床、哺乳、胚胎发育特点
常用实验动物着床、哺乳、胚胎发育特点可参考表10-42至10-44。
表10-42 各种动物着床时间
| 品 种 | 着 床 时 间 |
| 小 鼠 | 5天 |
| 大 鼠 | 5-6天 |
| 豚 鼠 | 6-7天 |
| 人 | 6-7天 |
| 兔 | 7-8天 |
| 雪 貂 | 7-8天 |
| 猕 猴 | 9-11天 |
| 猪 | 约11天 |
| 犬 | 13-14天 |
| 猫 | 13-14天 |
| 绵 羊 | 17-18天 |
| 牛 | 30-35天 |
| 马 | 8-9星期 |
表10-43 各种动物的哺乳期和乳腺数
| 动 物 | 哺 乳 期 | 乳 腺 数 |
| 狗 | 60天 | 5对 |
| 猫 | 7周(也有4~5周断奶的) | 4~6对 |
| 兔 子 | 42天 | 8~12对 |
| 豚 鼠 | 21天 | 1对 |
| 大白鼠 | 20~25天 | 6对(胸腺各3对) |
| 小白鼠 | 17~20天 | 5对(胸部3对腹部2对) |
| 田 鼠 | 21(20~25)天 | 7~8对 |
| 沙 鼠 | 21天 | |
| 老 鼠 | 20天 | |
| 猪 | 60天 | 5~7对 |
| 绵 羊 | 4个月 | 1对 |
| 山 羊 | 3个月 | 1对 |
表10-44 实验小鼠和大鼠各胚胎发育时期的变化
| 时期 | 胎龄(日) | 胚 胎 发 育 | |
| 小 鼠 | 大 鼠 | ||
| 1 | 1 | 1 | 单核细胞(输卵管内) |
| 2 | 2 | 2细胞期(输卵管内) | |
| 3 | 2 | 3.25 | 4细胞期(输卵管内) |
| 4 | 2.5 | 3.5 | 8-12细胞期(输卵管内) |
| 5 | 3 | 3. | 桑椹期(输卵管—子宫上部) |
| 6 | 3.5 | 4 | 囊胚初期(子宫内) |
| 7 | 4 | 5 | 囊胚期 |
| 8 | 4.5 | 6 | 植入开始 |
| 9 | 5 | 6.75 | 内外胚层分化 |
| 10 | 5.5 | 7.25 | 植入末期,卵黄腔形成,分化成内外胚层, |
| 11 | 6.5 | 7.75 | 植入完毕,出现羊膜。 |
| 12 | 7 | 8.5 | 出现中胚层和原条。 |
| 13 | 7.5 | 9 | 出现神经板,头部突起,出现尿膜褶。 |
| 14 | 7.75 | 9.5 | 体节1-4(头部)完成外层胎盘。 |
| 15 | 8~8.5 | 10 | 腔,胚外体腔内和羊膜腔发育。体节5-12(颈椎部) |
| 形成神经沟,胚回转开始。 | |||
| 16 | 8.5~9 | 10.5 | 体节13-20(胸椎前部)胚回转结束。 |
| 17 | 9.5 | 11 | 体节21-25(胸椎后部),神经沟闭锁。 |
| 18 | 10 | 11.5 | 体节26-28(腰椎前部),前肢出现。 |
| 19 | 10.25 | 11.75 | 体节29-31(腰椎后部),后肢出现。 |
| 20 | 体节32-33(荐椎后部)。 | ||
| 21 | 12 | 体节34、35(荐椎后部)。 | |
| 22 | 10.5 | 体节36(第一层尾椎)鼻窝形成 | |
| 23 | 体节37、38(尾椎)。 | ||
| 24 | 体节39、40(尾椎)。 | ||
| 25 | 11 | 12.5 | 体节41、42,前肢变大。 |
| 26 | 体节43-45(尾椎)手指分化。 | ||
| 27 | 12 | 13 | 体节46-48(尾椎)脐疝明显。 |
| 28 | 12.5 | 13.5 | 体节49-51(尾椎),鼻上颌闭锁。 |
| 29 | 14 | 体节52-55(尾椎)。 | |
| 30 | 13 | 14.5 | 体节56-60(尾椎),耳孔开,耳壳形成 |
| 31 | 13.5~14 | 15 | 体节61-63(尾椎),口盖板闭锁 |
| 32 | 14.5 | 15.5 | 体节64(尾椎)、体表出现毛囊。 |
| 33 | 15 | 16 | 体节65(最终尾椎) |
| 34 | 16-16.5 | 17-18 | 脐疝消失,眼脸、耳壳闭锁。 |
| 35 | 17-19 | 19-22 | 胎盘发育显著。临产。 |
(五)8常用实验动物的生殖结构特点
1.乳腺分布特点
常用实验动物乳腺分布特点见图10-9。
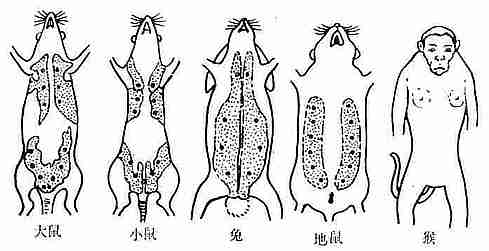
图10-9 常用实验动物乳腺分布特点
2.子宫结构特点
家兔的子宫属于双子宫型,是由一对几乎独立的、游离的子宫弯曲而成,两个子宫各有一个子宫颈分别与同一个阴道相通,即两个子宫的子宫颈共同开口于同一个阴道。所以,兔的子宫无所谓子宫角或子宫体,只是一对筒状结构,每只长度平均约7厘米,宽约3-4mm,但妊娠后期的子宫,在腹腔内可以扩展到很大的范围。
啮齿类动物的子宫均属双子宫型。其中,大白鼠的子宫呈Y字形排列,左左两个子宫颈开口于共同阴道和家兔的子宫一样,属于双子宫,双子宫颈,单阴道类型。
豚鼠的子宫属于双角型子宫,两个子宫角长约3~5cm,直径约5mm,背腹扁平。子宫体长约2cm,直径6mm,亦是背腹压扁。比子宫角略宽一点。子宫颈有两个内口,但只有一个公共外口。
肉食动物的子宫属于双角型子宫。具有发育良好的二个子宫角,单子宫体,单子宫颈和单阴道,不过左右两个子宫腔愈合范围较大,未愈合部分则形成弓状弯曲的子宫角。狗的子宫角细而长,子宫体很短,角体比例在5~6:1。中等体形的狗,子宫角长约12~15厘米,子宫体长约2~3厘米。另外,狗的子宫角腔内径均匀,没有弯曲,近乎直线。子宫颈很短,含有厚的肌肉层。
猫的双角子宫也很明显,但子宫体约4厘米,无论是相对比例还是绝对长度均比狗的子宫体长。
灵长类的子宫属于单子宫型、梨状,无子宫角,输卵管直接开口于子宫腔。常用实验动物子宫类型见图10-10。
3.胎盘类型牛、马、猪为上皮绒膜型;绵羊、山羊为结缔绒膜型;狗、猫为内皮绒膜型;兔为血内皮型;大鼠、豚鼠、恒河猴为血绒膜型。
4.生殖腺结构 可参看表10-45和10-46。
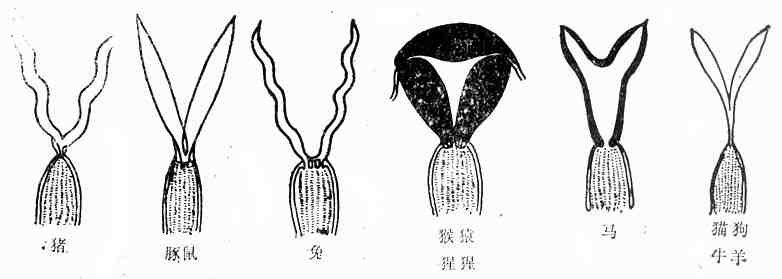
图10-10 常用实验动物子宫类型
表10-45 雄性动物生殖结构的比较解剖特点
| 动物种类 | 睾丸 | 壶腹腺 | 储精囊 | 前列腺 | 尿道球部腺体 | 前列周围腺体 | 阴茎骨 | 尿道腺体 | 球腺体 | 包皮腺体 | 腹股沟腺体 |
| 蝙蝠 | 3×2 | 4×15 | A | 5×3 | 1×1 | A | F | F | A | A | A |
| 猫 | 14×8 | A | A | 5×2 | 4×3 | A | F | A | A | A | A |
| 狗 | 40×30 | F | A | 25×16 | A | A | F | A | A | A | A |
| 沙土鼠 | 14×9 | 6×5 | 20×12 | 9×7 | 6×4 | A | F | F | A | A | A |
| 豚鼠 | 25×15 | A | 115×7 | 15×8 | 8×5 | A | F | A | A | PD | A |
| 田鼠 | 14×11 | 3×3 | 11×6 | 8×6 | 4×3 | A | F | F | A | 3×1 | A |
| 貂类 | 11×8 | F | A | 10×5 | A | A | F | A | A | A | A |
| 小鼠 | 6×4 | 2×1 | 13×4 | 4×4 | 3×2 | A | F | F | A | 6×5 | A |
| 兔(美国) | 35×15 | 15×4 | 19×7 | 19×6 | 6×3 | 6×2 | A | A | A | A | 15×7 |
| 袋鼠 | 15×11 | A | A | 40×9 | 19×15 | A | A | A | A | A | A |
| 兔 | 35×17 | F | A | 25×6 | 6×2 | A | A | A | A | A | 10×4 |
| 大鼠 | 20×12 | 4×4 | 20×10 | 13×10 | 5×3 | A | F | F | A | 16×4 | A |
| 地鼠 | 3×2 | 3×1 | A | 3×1.5 | 1×1 | A | A | A | A | 分散的 | A |
| 树松鼠 | 30×12 | F | 7×4 | 28×18 | 13×13 | A | F | A | 10×7 | A | A |
| 地松鼠 | 14×8 | F | 8×7 | 18×18 | 8×6 | A | F | A | 10×8 | F | A |
| 类人猿 | 10×7 | A | 14×7 | 6×5 | A | A | F | A | A | 分散的 | A |
| 雪貂 | F | F | A | F | A | A | F | A | A | A |
注①F表示属于一种功能性腺体不能估量
②A表示这种结构不发生
③PD表示这种腺体是不发达的
④测量的单位为毫米
表10-46实验动物的副性腺
| 动 物 | 精 囊 | 前 列 腺 | 尿道球腺 | 凝 固 腺 | 包 皮 腺 |
| 狗 | - | √ | - | - | - |
| 猫 | - | √ | √ | - | - |
| 兔 | √ | √ | √ | - | - |
| 豚鼠 | √ | √ | √ | - | - |
| 鼠类(大、小鼠) | √ | √ | √ | √ | √ |
| 猪 | √ | √ | √ | - | - |
| 羊 | √ | √ | √ | - | - |
狗缺少精囊和尿道球道腺,猫也没有精囊,雄兔还有前列旁腺,鼠类多一种凝固腺,大鼠和小鼠有包皮腺。
二、动物的选择与应用
(一)口服避孕药物的筛选研究
目前国内外广泛应用的口服避孕药(孕激素与雌激素的复合片以及单纯孕激素药片)其避孕效果都非常好,但还存在一定副作用。为了提高口服避孕药的可接受率,有必要寻找更加理想的、有效、安全、经济、方便的药物。
筛选口服避孕药的实验动物多使用啮齿类实验动物,因为它们具有规律的动情周期,排卵有明显的指标,易于检测,价格便宜,而具有月经周期的灵长类动物反而不是都被选择的对象。因为这类动物来源困难,价格昂贵,排卵和月经周期难于检测。
当动物实验证实某药物有抗生育的效果后,还必须进行各种药理实验。
1.孕激素活性测定 根据Mcphail的方法,以未成熟雌性家兔子宫内膜的变化为观察指标;
2.妊娠维持实验 一般选大鼠为动物模型;
3.雌激素活性及抗雌激素活性测定 用未成年或去卵巢未成熟大鼠为动物模型,以子宫称重或阴道涂片为观察指标;
4.雄激素活性及抗雄激素活性测定 用大鼠或小鼠为动物模型,以前列腺腹侧及储精囊称重为观察指标;
5.对促性腺激素分泌的影响 用啮齿类或其它种类动物为实验模型,以是否抑制排卵或测定外周血的促性腺激素含量为观察指标;
6.蜕镇痛形成实验 用小鼠为动物模型;
7.糖皮质激素活性测定 用大量为动物模型。
以上7个为基本实验,根据需要还可以增添其它实验。
(二)避孕药物的药理研究
1. 女用避孕药物 研究最基本方法是抗生育实验,就是在与雄性动物合笼前和合笼交配期间,给雌性动物服药,观察药物对生育的影响。以检查雌性动物怀孕百分率和每只孕鼠之胚胎数为指标。如果能找到一种药具有防止受精卵着床的作用或中断早期妊娠的作用,那就不必天天服药,而只在需要时服用。这是避孕药研究工作者多年来的一项设想。常需要作抗着床和抗早孕实验。抗着床实验是将雌鼠与雄鼠合笼,每天上午检查雌鼠阴道口有无阴栓,于出现阴栓后第1~5天,每日给药一次,于阴栓出现后第12天处死动物,以胚胎数和怀孕百分率为指标。阴栓是某些啮齿类动物在交配后,随即形成的一种白色浆液性物质,堵塞在阴道至子宫颈的腔内,称为阴栓(Vaginal plug)。是检查动物已否交配的一种标志,将出现阴栓的当天算为妊娠“零天”。阴栓是由雄鼠精囊腺和凝固腺的分泌物在阴道内凝固而成的产物。小白鼠一般在交配后4~5分钟内形成。大的阴栓可凸出于阴道口,小的阴道或子宫颈内,需要翻开阴道口才能查见。阴栓一般在交配后一小时起渐渐软化,经16~24小时后脱落。但也有只存在4~5小时,而最多的可存在48小时以上。小鼠阴栓比较牢固,大鼠的阴栓不牢固,容易脱落,所以检查大鼠阴栓时,除检查阴道外,还应在笼底寻找阴栓。亦可用生理盐水冲洗阴道,在显微镜下检查冲洗液中有无精子,有精子表示已交配。抗早孕实验是在着床(出现阴栓后第5天着床)以后给药,一般于阴栓出现后第6~9天连续给药4天,第12天处理,以检查胚胎数和死胎为指标。抗生育、抗着床和抗早孕实验中最常用的动物为壮年的28~35g的小鼠,也可用体重200~250体重的大鼠。在探讨一个避孕药的作用原理时,有时需要检查一个药物有没有抑制排卵作用。需要进行抗排卵实验,常选用体重2~3.5kg的雌兔进行实验。
2.男用避孕药物研究 长期以来,对男用避孕药的研究不很重现,进展缓慢。药物能抑制精子的生成过程,干扰男性生殖活动的激素调节;干扰副性腺和改变精液的理化特性等,均可达到不孕的目的。现在认为,精子形态的成熟是在睾丸,而生理的成熟是在副睾。药物作用于副性腺,干扰精子成熟或改变精液理化性质所引起的男性不孕似较理想。常选用成年雄性大白鼠作实验,如观察药物对大白鼠生育力的影响;对大白鼠精子数的影响;对大白鼠精子活动力的影响以及对雄性激素的作用等。
实验中常需要采集实验动物的精液,采集的方法有电刺激法、假阴道法、按摩法以及直接杀取法。电刺激法适用于多种实验动物,尤其是野生实验动物的采精,其原理是通过刺激输卵管壶腹附近的神经末稍,引起低级射精中枢兴奋而促使动物射精。所用电刺激采精器的主要技术参数为:电源电压-交流220V;输出可调电压0~20V;可高频率20~60Hz:输出电流0~1000mA:输出波形-正弦波或方波。假阴道法是用硬质外壳衬以胶质内胎制成的采精器械,内部注以温水和充气,适当加以润滑,使其与雌性动物生殖道的内环境相似,以诱起雄性动物射精常,常适用于兔的采精。猫在有发情母猫引诱情况下,当公猫爬跨时,也可使用此法采精,但象水貂那样在交配时雌雄动物长时间紧密拥抱的动物,无法使用假阴道。按摩法主要用于狗的采精,这是因为狗在交配时,雄狗阴茎的球状海绵体充血胀大并和雌狗的阴道锁结,采取假阴道法采精有一定困难,故多采用按摩法。狗的射精分为3段,前段为尿道腺的分泌物,约为0.25~2.0ml的透明液体:中段排出的是浓精,约0.5~3.0ml;第3段则为大量透明的前列腺分泌物,量可达3~20ml,但几乎不含精子。按摩法对于调教好的雄狗方便易行,并可存细胞观察射精情况,收集所需部分,无需特殊设备。但不能用于野生犬科或陌生猛犬。直接杀取法适于前面几种方法不适用时采用,如小白鼠、水貂等,系将动物处死,取其输卵管和附睾丸,用生理盐水或其它专门稀释液冲洗,制成精子悬浮液备用。常用实验动物采精方法和精液特点见表10-47。
表10-47 各种动物精液采取方法和精液特点
| 动物 | 采取法 | 交配时间 | 射精时间 | 颜色 | 精液量(毫升) | 精液密度万/立方毫米 | 活力 | 畸形率% | pH |
| 牛 | 假阴道法电刺激法 | 瞬间 | 瞬间 | 乳白 | 5~6 | 90~100 | 卅70< | 10 | 6.6~6.8 |
| 马 | 假阴道法阴道内采取 | 暂时 | 数秒至10数秒 | 淡 | 50~150 | 20~30 | 同上 | 5 | 7.2~7.6 |
| 猪 | 假阴道法 | 10分 | 5~10分 | 灰白 | 200~300 | 20 | 同上 | 5~10 | 6.4~7.4 |
| 绵羊 | 假阴道法电刺激法 | 瞬间 | 瞬间 | 乳白 | 0.5~0.2 | 300 | 同上 | 同上 | 6.4~7.2 |
| 山羊 | 同上 | 同上 | 同上 | 乳白带黄 | 同上 | 同上 | 同上 | 同上 | 同上 |
| 狗 | 按摩法 假阴道法 | 10分 | 1分 | 乳白 | 0.5~3.0 | 10~15 | 同上 | 10~15 | 6.6~6.8 |
| 家兔 | 假阴道法阴道内采取 | 10秒 | 瞬间 | 乳白 | 0.5~1.0 | 50~60 | 同上 | 同上 | 6.6~6.8 |
| 大鼠 | 乙醚麻醉 | 同上 | 同上 | 乳白 | 同上 | 0.1 | 同上 | - | |
| 鸡 | 电刺激法 | 2秒 | 1秒 | 同上 | 0.1~0.5 | 200~500 | 卄卅 70< | <20 | 6.5~7.4 |
(三)中枢神经系统对促性腺激素分泌的调节作用研究
不同动物的中枢神经系统对垂体促性腺激素分泌的调节是有很大差异的。如灵长类和大鼠的垂体以同样的方式对促黄体素释放激素起反应,并以同样的形式被大剂量雌激素或雄激素所抑制。这两种动物的中枢神经系统,在自发性排卵中都是必不可少的,这是它们的共同点。但调节大鼠促性腺激素释放的中枢是视前结节系统,若视前区受损,则在排卵前雌激素不能导致促黄体激素的释放。而这一区域受损不影响恒河猴促黄体激素的释放,因为恒河猴促性腺素释放的调节中枢是结节漏头斗状系统,这是它们的不同点。
在中枢神经系统对促性腺激素调节的研究中,大鼠是常用的动物模型,因这方面已积累了大量的有关研究资料:价格便宜,易于饲养,有规律的性周期便于检测,且周期短,易于安排实验,排卵前有自发的LH峰,所得的结果一般易于在猴子身上得到重复。
阴道涂片可以准确地判断大鼠和小鼠动情周期的各个阶段,从而可推测排卵前LH峰及排卵,但此方法很少用于田鼠,因为田鼠在动情周期阴道有渗出物,所以不一定要做阴道涂片。推测灵长类LH峰,须测定外周血的孕激素水平,如单独根据月经出血期来推测有可能导致判断的错误。
大鼠、小鼠和田鼠的黄体期不能自发地变成功能性黄体,在正常情况下,需交配诱发黄体激素的分泌,而恒河猴和人则自发性功能黄体。家兔和猫是诱发性(交配)排卵,一旦排卵发生,则黄体能自发地变为功能性的。
(四)神经内分泌学研究
通过对人性行为的内分泌研究,发现人类性活动大多可以用动物实验来阐明,而最好的动物模型不一定是灵长类。Nadle分析了大猩猩、黑猩猩和猩猩的性活动行为,结果发现在新皮层发育及智能上,他们大体上是相等的,但在性活动方面却有很大的不同,大猩猩交配大多集中在月经周期中期,且雌性动物起主导作用,猩猩可以在周期的任何时期进行交配,没有明显的排卵标志,雄性动物的性要求很强烈,在交配中占主导地位,黑猩猩属这两者之间。所以虽然灵长类最接近人类,但用它作神经内分泌学研究所得的结果也能完全适用于人,而较小的动物如大鼠,其性活动与人也有共同点,如在大鼠的动物实验及人类的临床观察中,均存在维持性功能的雄性激素水平比维持生育功能的水平低,人类性功能随年龄的增长而减退,某些器官出现肥大等变化在大鼠也都有发生,所以大鼠用于性活动研究,也是一个较好的模型。