第二节 嘌呤核苷酸的合成代谢
一、嘌呤核苷酸的合成
体内嘌呤核苷酸的合成有两条途径:①利用磷酸核糖、氨基酸、一碳单位及CO2等简单物质为原料合成嘌呤核苷酸的过程,称为从头合成途径(denovo synthesis),是体内的主要合成途径。②利用体内游离嘌呤或嘌呤核苷,经简单反应过程生成嘌呤核苷酸的过程,称重新利用(或补救合成)途径(saluagepathway)。在部分组织如脑、骨髓中只能通过此途径合成核苷酸。
(一)嘌呤核苷酸的从头合成
早在1948年,Buchanan等采用同位素标记不同化合物喂养鸽子,并测定排出的尿酸中标记原子的位置的同位素示踪技术,证实合成嘌呤的前身物为:氨基酸(甘氨酸、天门冬氨酸、和谷氨酰胺)、CO2和一碳单位(N10甲酰FH4,N、N10-甲炔FH4)(图8-3)。
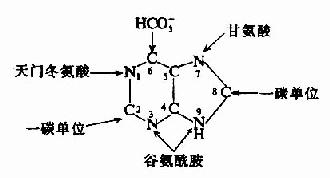
图8-3 嘌呤环合成的原料来源
随后,由Buchanan和Greenberg等进一步搞清了嘌呤核苷酸的合成过程。出人意料的是,体内嘌呤核苷酸的合成并非先合成嘌呤碱基,然后再与核糖及磷酸结合,而是在磷酸核糖的基础上逐步合成嘌呤核苷酸。嘌呤核苷酸的从头合成主要在胞液中进行,可分为两个阶段:首先合成次黄嘌呤核苷酸(inosine monophosphate IMP);然后通过不同途径分别生成AMP和GMP。下面分步介绍嘌呤核苷酸的合成过程。
1.IMP的合成:IMP的合成包括11步反应:(图8-4)
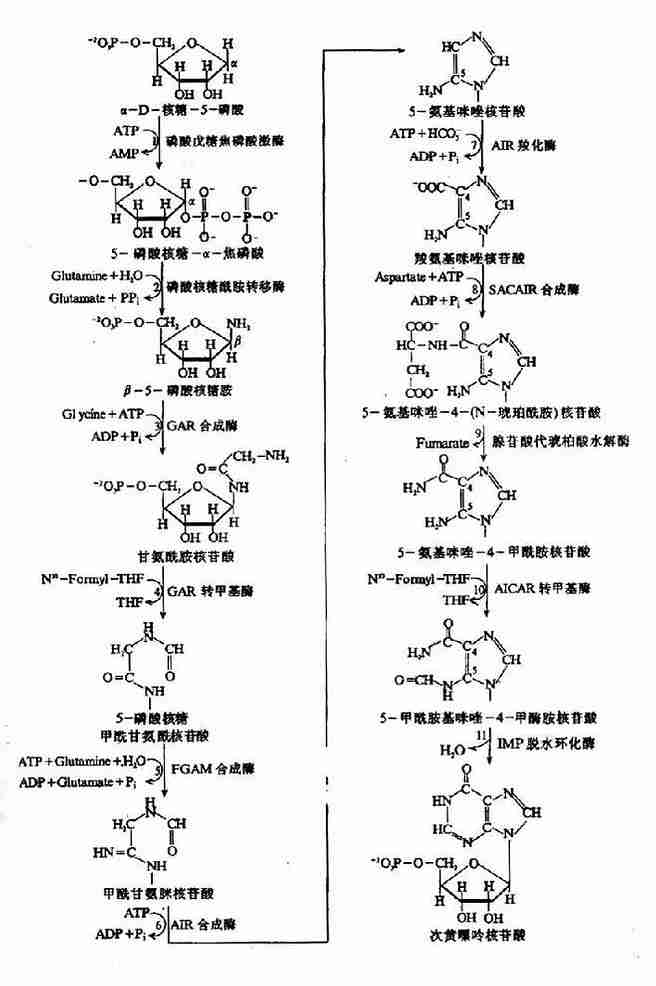
图8-4 IMP的合成
(1)5-磷酸核糖的活化:嘌呤核苷酸合成的起始物为α-D-核糖-5-磷酸,是磷酸戊糖途径代谢产物。嘌呤核苷酸生物合成的第一步是由磷酸戊糖焦磷酸激酶(ribosephosphate pyrophosphohinase)催化,与ATP反应生成5-磷酸核糖-α-焦磷酸(5-phosphorlbosyl?α-pyrophosphatePRPP)。此反应中ATP的焦磷酸根直接转移到5-磷酸核糖C1位上。PRPP同时也是嘧啶核苷酸及组氨酸、色氨酸合成的前体。因此,磷酸戊糖焦磷酸激酶是多种生物合成过程的重要酶,此酶为一变构酶,受多种代谢产物的变构调节。如PPi和2,3-DPG为其变构激活剂。ADP和GDP为变构抑制剂。
(2)获得嘌呤的N9原子:由磷酸核糖酰胺转移酶(amidophosphoribosyltransterase)催化,谷氨酰胺提供酰胺基取代PRPP的焦磷酸基团,形成β-5-磷酸核糖胺(β-5-phosphoribasylamine PRA)。此步反应由焦磷酸的水解供能,是嘌呤合成的限速步骤。酰胺转移酶为限速酶,受嘌呤核苷酸的反馈抑制。
(3)获得嘌呤C4、C5和N7原子:由甘氨酰胺核苷酸合成酶(glycinamide ribotidesynthetase)催化甘氨酸与PRA缩合,生成甘氨酰胺核苷酸(glycinamide ribotide,GAR)。由ATP水解供能。此步反应为可逆反应,是合成过程中唯一可同时获得多个原子的反应。
(4)获得嘌呤C8原子:GAR的自由α-氨基甲酰化生成甲酰甘氨酰胺核苷酸(formylgly?cinamide ribotideFGAR)。由N10-甲酰-FH4提供甲酰基。催化此反应的酶为GAR甲酰转移酶(GAr transtormylase)。
(5)获得嘌呤的N3原子:第二个谷氨酰胺的酰胺基转移到正在生成的嘌呤环上,生成甲酰甘氨脒核苷酸(formylglycinamidine ribotide,FGAM)。此反应为耗能反应,由ATP水解生成ADP+Pi,供能。
(6)嘌呤咪唑环的形成:FGAM经过耗能的分子内重排,环化生成5-氨基咪唑核苷酸(5-aminoimidazole ribotide,AIR)。
(7)获得嘌呤C6原子:C6原子由CO2提供,由AIR羧化酶(AIrcarboxylase)催化生成羧基氨基咪唑核苷酸(carboxyamino imidazoleribotide,CAIR)。
(8)获得N1原子:由天门冬氨酸与AIR缩合反应,生成5-氨基咪唑-4-(N-琥珀酰胺)核苷酸(4-aminoimidazole-4-(N-succinylocarboxamide)ribotide,SACAIR)。此反应与(3)步相似,由ATP水解供能。
(9)去除延胡索酸:SACAIR在SACAIR甲酰转移酶催化下脱去延胡索酸生成5-氨基咪唑-4-甲酰胺核苷酸(5-aminoimidazole-4-carboxamideribotide,AICAR)。(8)、(9)两步反应与尿素循环中精氨酸生成鸟氨酸的反应相似。
(10)获得C2:嘌呤环的最后一个C原子由N10-甲酰-FH4提供,由AICAR甲酰转移酶催化AICAR甲酰化生成5-甲酰胺基咪唑-4-甲酰胺核苷酸(5-formaminoimidazole-4carboxyamideribotide,FAICAR)。
(11)环化生成IMP:FAICAR脱水环化生成 IMP。与反应(6)相反,此环化反应无需ATP供能。
2.由IMP生成AMP和GMP
上述反应生成的IMP并不堆积在细胞内,而是迅速转变为AMP和GMP。AMP与IMP的差别仅是6位酮基被氨基取代(图8-5)。此反应由两步反应完成。(1)天门冬氨酸的氨基与IMP相连生成腺苷酸代琥珀酸(adenylosuccinate),由腺苷酸代琥珀酸合成酶催化,GTP水解供能。(2)在腺苷酸代琥珀酸裂解酶作用下脱去延胡索酸生成AMP。
GMP的生成也由二步反应完成。(1)IMP由IMP脱氢酶催化,以NAD+为受氢体,氧化生成黄嘌呤核苷酸(xanthosinemonophosphate,XMP)。(2)谷氨酰胺提供酰胺基取代XMP中C2上的氧生成GMP,此反应由GMP合成酶催化,由ATP水解供能(图8-5)。
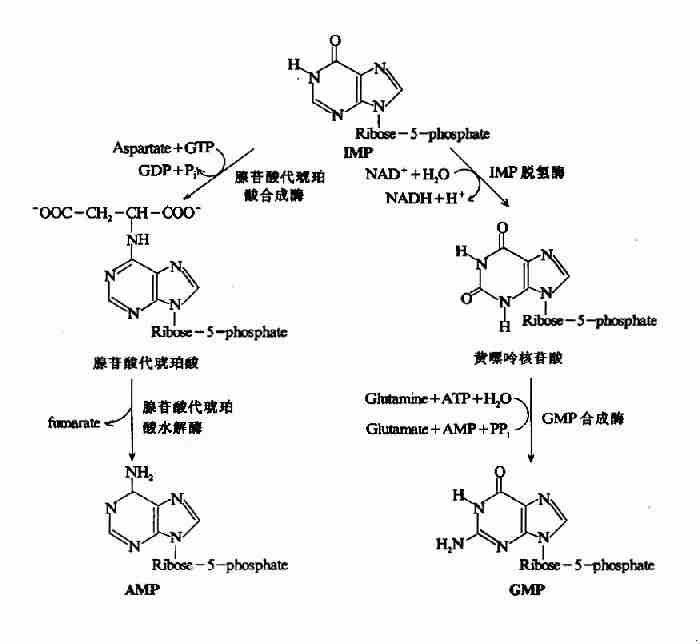
图8-5 IMP分别生成AMP和GMP
3.一磷酸核苷磷酸化生成二磷酸核苷和三磷核苷。
要参与核酸的合成。一磷酸核苷必须先转变为二磷酸核苷再进一步转变为三磷酸核苷。二磷酸核苷由碱基特异的核苷一磷酸激酶(nucleoside monophosphate kinase)催化,由相应一磷酸核苷生成。例如腺苷激酶催化AMP磷酸化生成ADP

二磷酸核苷激酶对底物的碱基及戊糖(核糖或脱氧核糖)均无特异性。此酶催化反应系通过“乒乓机制”,即底物NTP使酶分子的组氨酶残基磷酸化,进而催化底物NDP的磷酸化。反应△G≈0,为可逆反应。
4.嘌呤核苷酸从头合成的调节
从头合成是体内合成嘌呤核苷酸的主要途径。但此过程要消耗氨基酸及ATP。机体对合成速度有着精细的调节。在大多数细胞中,分别调节IMP,ATP和GTP的合成,不仅调节嘌呤核苷酸的总量,而且使ATP和GTP的水平保持相对平衡。嘌呤核苷酸合成调节网可见图8-6。
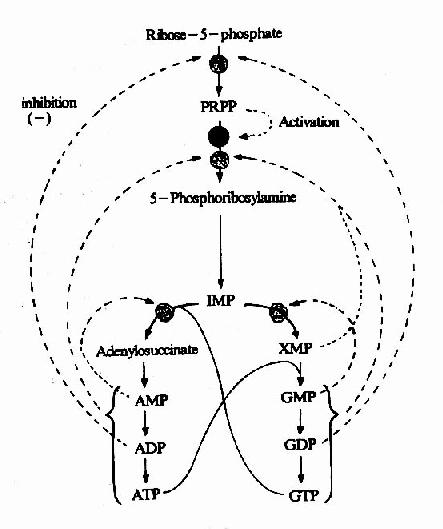
图8-6 嘌呤合成的调节网
IMP途径的调节主要在合成的前二步反应,即催化PRPP和PRA的生成。核糖磷酸焦磷酸激酶受ADP和GDP的反馈抑制。磷酸核糖酰胺转移酶受到ATP、ADP、AMP及GTP、GDP、GMP的反馈抑制。ATP、ADP和AMP结合酶的一个抑制位点,而GTP、GDP和GMP结合另一抑制位点。因此,IMP的生成速率受腺嘌呤和鸟嘌呤核苷酸的独立和协同调节。此外,PRPP可变构激活磷酸核糖酰胺转移酶。
第二水平的调节作用于IMP向AMP和GMP转变过程。GMP反馈抑制IMP向XMP转变,AMP则反馈抑制IMP转变为腺苷酸代琥珀酸,从而防正生成过多AMP和GMP。此外,腺嘌呤和鸟嘌呤的合成是平衡。GTP加速IMP向AMP转变,而ATP则可促进GMP的生成,这样使腺嘌呤和鸟嘌呤核苷酸的水平保持相对平衡,以满足核酸合成的需要。
(二)补救合成途径:
大多数细胞更新其核酸(尤其是RNA)过程中,要分解核酸产生核苷和游离碱基。细胞利用游离碱基或核苷重新合成相应核苷酸的过程称为补救合成(saluage pathway)。与从头合成不同,补救合成过程较简单,消耗能量亦较少。由二种特异性不同的酶参与嘌呤核苷酸的补救合成。腺嘌呤磷酸核糖转移酶(Adenine phosphoribosyl transterase,APRT)催化PRPP与腺嘌呤合成AMP:


人体由嘌呤核苷的补救合成只能通过腺苷激酶催化,使腺嘌呤核苷生成腺嘌呤核苷酸。
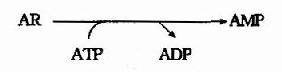
嘌呤核苷酸补救合成是一种次要途径。其生理意义一方面在于可以节省能量及减少氨基酸的消耗。另一方面对某些缺乏主要合成途径的组织,如人的白细胞和血小板、脑、骨髓、脾等,具有重要的生理意义。例如Seseh?Nyhan综合征是由于HGPRT的严重遗传缺陷所致。此种疾病是一种性?连锁遗传缺陷,见于男性。患者表现为尿酸增高及神经异常。如脑发育不全、智力低下、攻击和破坏性行为、常咬伤自己的嘴唇、手和足趾,故亦称自毁容貌症。其尿酸增高较易解释,由于HGPRT缺乏,使得分解产生的PRPP不能被利用而堆积,PRPP促进嘌呤的从头合成,从而使嘌呤分解产物-尿酸增高。而神经系统症状的机制尚不清楚。